Archives
- 2018-07
- 2019-04
- 2019-05
- 2019-06
- 2019-07
- 2019-08
- 2019-09
- 2019-10
- 2019-11
- 2019-12
- 2020-01
- 2020-02
- 2020-03
- 2020-04
- 2020-05
- 2020-06
- 2020-07
- 2020-08
- 2020-09
- 2020-10
- 2020-11
- 2020-12
- 2021-01
- 2021-02
- 2021-03
- 2021-04
- 2021-05
- 2021-06
- 2021-07
- 2021-08
- 2021-09
- 2021-10
- 2021-11
- 2021-12
- 2022-01
- 2022-02
- 2022-03
- 2022-04
- 2022-05
- 2022-06
- 2022-07
- 2022-08
- 2022-09
- 2022-10
- 2022-11
- 2022-12
- 2023-01
- 2023-02
- 2023-03
- 2023-04
- 2023-05
- 2023-06
- 2023-07
- 2023-08
- 2023-09
- 2023-10
- 2023-11
- 2023-12
- 2024-01
- 2024-02
- 2024-03
- 2024-04
- 2024-05
- 2024-06
- 2024-07
- 2024-08
- 2024-09
- 2024-10
- 2024-11
- 2024-12
- 2025-01
- 2025-02
- 2025-03
-
br Introduction Myotonic dystrophy dystrophia myotonica DM i
2020-06-12
Introduction Myotonic dystrophy (dystrophia myotonica, DM) is an autosomal dominant disorder and the most common form of inherited muscular dystrophy in adults [1]. DM is characterised by a wide range of symptoms, including myotonia, progressive muscle loss, cataracts, cardiac conduction defects,
-
Given that dimethylamine substitution was tolerated as in SR
2020-06-12
Given that dimethylamine substitution was tolerated as in SR106447 (), we investigated nmda receptor amine substitutions as replacements for the -butyl group (). Cyclic amines (SR19880, 19882) or simple mono-substituted anilines (SR20037) were not active, nor was an acetylated version (SR20039). Th
-
TTNPB br Author Contributions br Acknowledgments
2020-06-12

Author Contributions Acknowledgments This work was supported, in part, by a grant from the National Cancer Institute of the National Institutes of Health under Grant U01CA202241 and National Institutes of Health under Grant R01NS083856. Introduction The Eph (erythropoietin-producing human
-
Several effective strategies have been developed to design a
2020-06-12
Several effective strategies have been developed to design activatable MR probes according to the Solomon, Bloembergen and Morgan (SBM) theory, including modulation of the number of inner-sphere water molecules (q), the rotational tumbling time (τ) and the residence lifetime of inner-sphere water mo
-
br Materials and methods br Results br Discussion Endothelia
2020-06-12
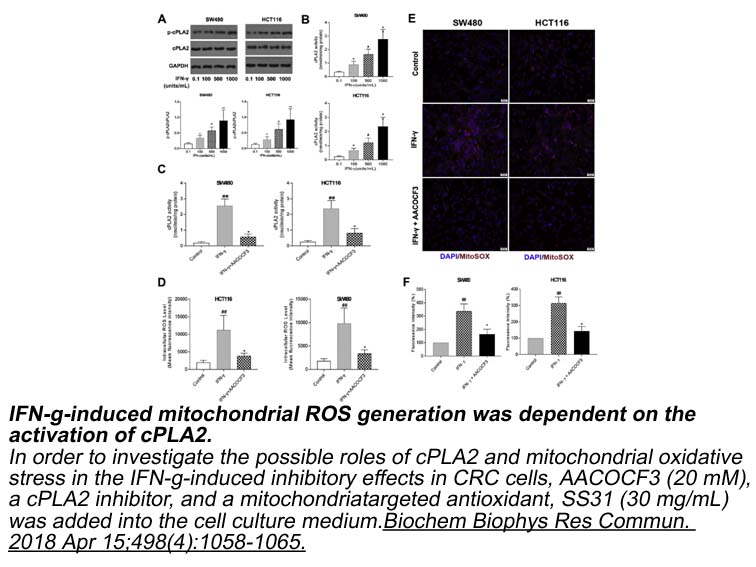
Materials and methods Results Discussion Endothelial cells are crucial for both vascular homeostasis and protecting the vasculature against atherogenic insults [3]. OxLDL-mediated injury to endothelial cells is crucial for endothelial dysfunction in the pathogenesis of atherosclerosis and a
-
MicroRNA miRNA is a class of
2020-06-12
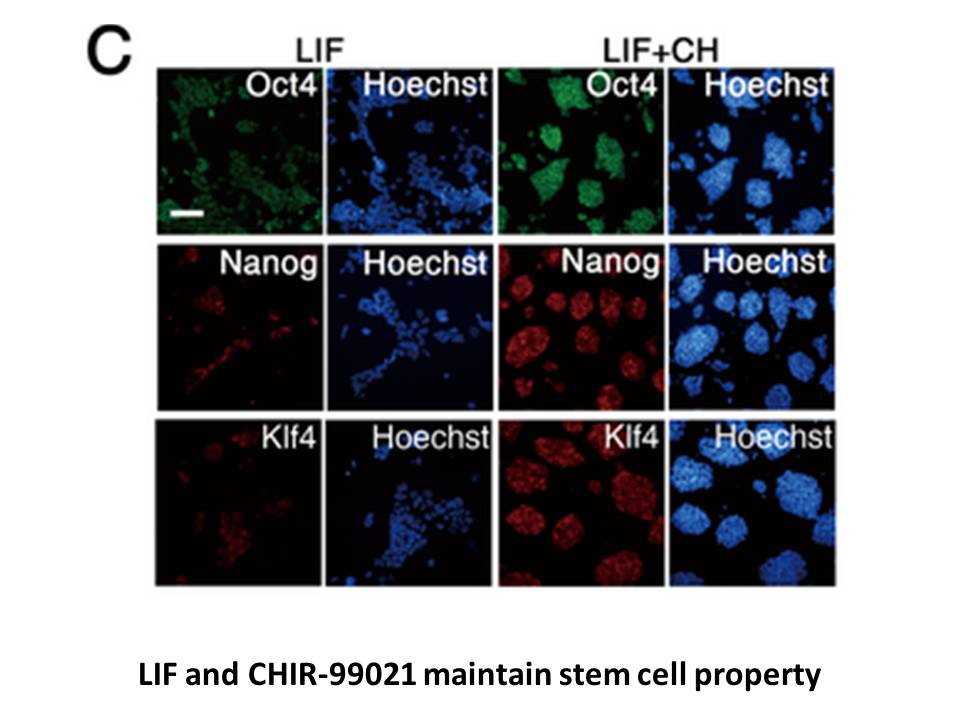
MicroRNA (miRNA) is a class of highly conserved, non-coding small molecules containing 19–25 nucleotides. miRNAs are involved in the regulation of genes associated with cancer development and progression and therefore can act as oncogenes, by contributing to tumor formation,17, 18 or as tumor suppre
-
The mechanism by which these HIV PIs impair skeletal
2020-06-12
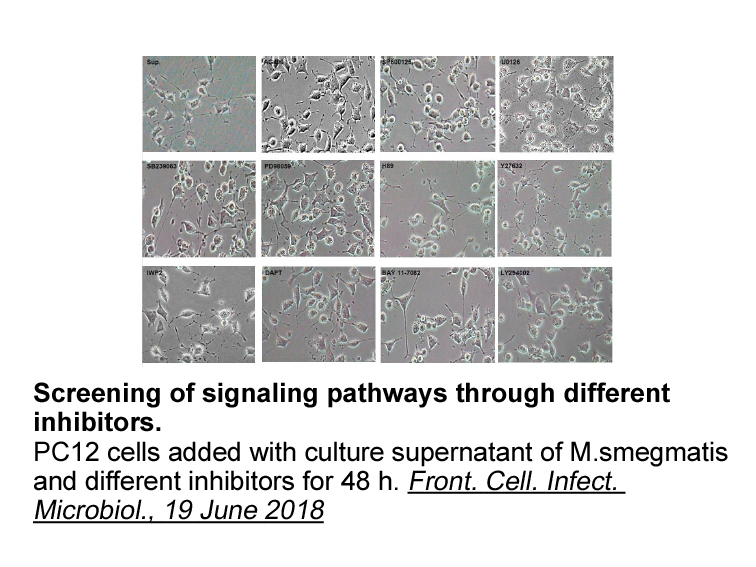
The mechanism by which these HIV-PIs impair skeletal muscle palmitate transport and oxidation has been partially elucidated. CD36 (also referred to as fatty PHA-848125 translocase; FAT) is a transmembrane protein involved in the transport of long-chain fatty acids (LCFA) across cellular membranes.
-
As expected the nortestosterone derived progestins tested we
2020-06-12

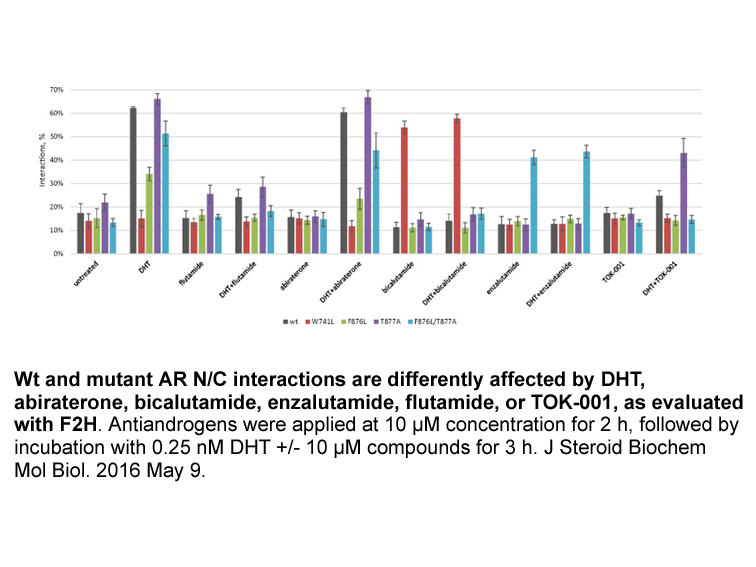
As expected, the 19-nortestosterone-derived progestins tested were moderate agonists of hAR (C), with EC50s ranging from 2 to 10nM, however all were less potent than the endogenous ligand DHT by at least a factor of 6, with relative potencies ranging from 0.0301 to 0.164. This is in agreement with p
-
AAG has a broad substrate
2020-06-11
AAG has a broad substrate specificity and besides 3-methyladenine can excise other altered purine residues, such as the minor lesions hypoxanthine and 1, N6-ethenoadenine from DNA. AAG initially activates these neutral base lesions by protonation of the base to allow for general DC_AC50 catalysis [
-
Previous and ongoing clinical trials
2020-06-11

Previous and ongoing clinical trials have been summarized in a recent review (Siaw-Debrah et al., 2017). To our knowledge, there are no ongoing or completed clinical trials focusing on cytokines in ICH. Published studies have focused mainly on cytokine changes in ICH patients (summarized in Table 3)
-
orbifloxacin Phylogenetic analysis indicates that in poikilo
2020-06-11
Phylogenetic analysis indicates that in poikilothermic vertebrates CXCR3a and CXCR3b have evolved independently from a common gene possibly after the 2nd round whole genome duplication but before the split of bony fish and tetrapods (Fig. 1 and Supplementary file 3). Owing to the extra round of whol
-
br Limitations of the study br Possible conflicts of interes
2020-06-11
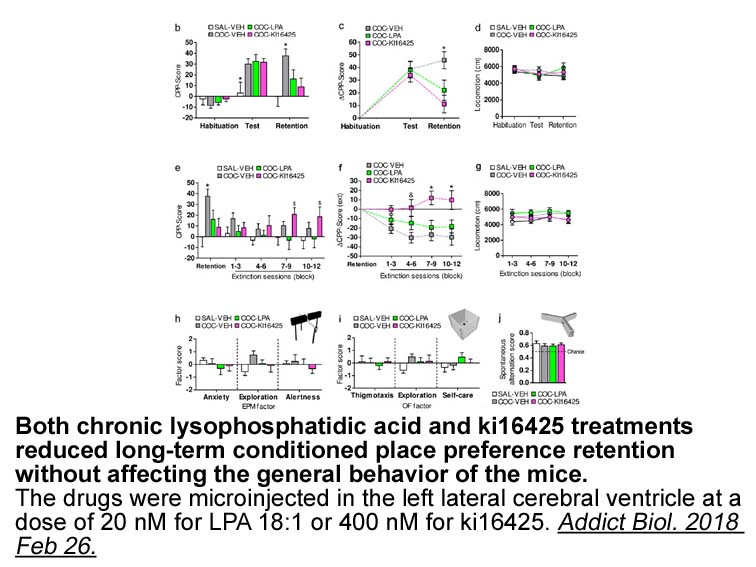
Limitations of the study Possible conflicts of interest Dr. Fayemiwo or SAF has received full financial support from Europe Gilead Sciences Ltd. for his M.Sc. degree programme in Medical Mycology and has been paid for talks on behalf of AstraZeneca and GSK. Caroline Moore or CBM has received
-
DAPI hydrochloride Quantum walks can be connected to extende
2020-06-11

Quantum walks can be connected to extended CMV matrices as follows. If all Verblunsky coefficients with even index vanish, then the extended CMV matrix becomes which resembles the matrix representation U. One may notice, however, that in the extended CMV matrix , whereas in the quantum walk matrix
-
EphB promotes tumor growth by stimulating angiogenesis throu
2020-06-11
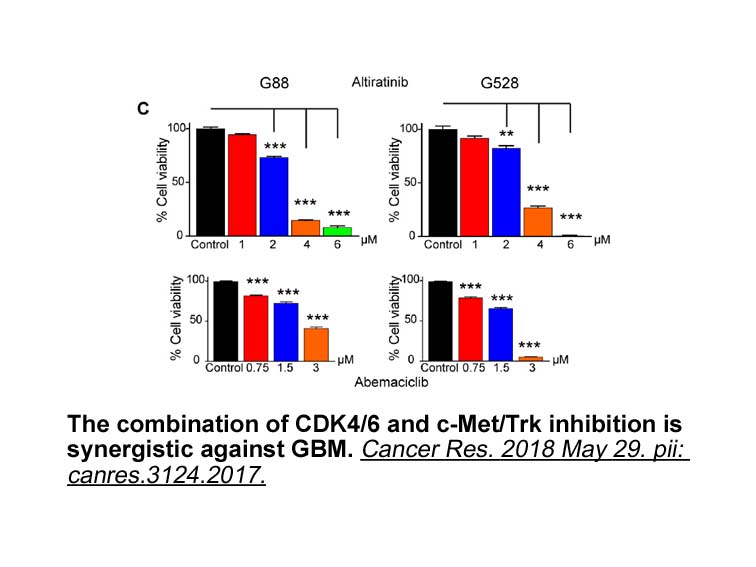
EphB4 promotes tumor growth by stimulating angiogenesis through EphrinB2 [18]. However, the distinct and specifically involvement of AMD-070 australia and veins in the tumor angiogenesis of all tumor grades is unclear. Our results indicate that the relation between EphrinB2 and EphB4 stained blood
-
Grapiprant is a selective antagonist
2020-06-11
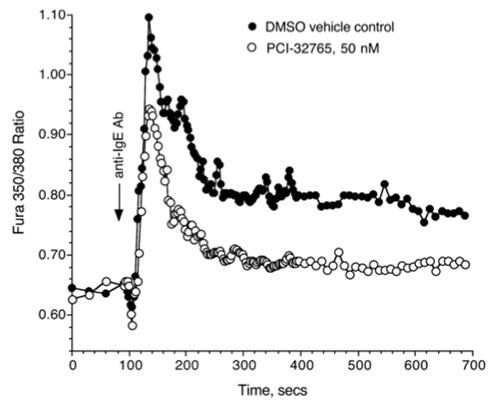
Grapiprant () is a selective antagonist for prostaglandin E (PGE) receptor subtype 4 (EP4) identified as a clinical candidate for the treatment of inflammatory pain associated with osteoarthritis (OA). It is currently under development for use in humans and dogs. The projected dosing regimen of grap
15357 records 800/1024 page Previous Next First page 上5页 796797798799800 下5页 Last page